A new study out of China is sounding the alarm over a new phenomenon called “quarantine myopia”, which experts say could have a major long-term impact on young children’s eyesight.
Myopia is also known as near-sightedness, and it can worsen and cause major problems as time goes on. Researchers say more severe outcomes, including loss of vision, are more likely the younger a child is when myopia first develops.
The study was published in JAMA Ophthalmology, according to CTV News, and looks into the amount of children in China who developed near-sightedness after lockdowns.
Researchers say the amount of children with myopia increased significantly in all age groups in 2020, more so than in any other year. It was especially prevalent with children between 6 and 8 years old.
The prevalence of myopia in kids aged 6 to 13 jumped by 10 to 15 per cent in 2020 compared to 2019.
An expert told CTV News that the problem isn’t necessarily screens themselves, it’s doing anything that requires focusing on something very near to the face for a very long time.
She also said the impacts were certainly the combination of a lack of outdoor activities, and increased screen time through online learning during the lockdown.
As for solutions, one possibility is for every 20 minutes of “near work”, like reading or looking at a computer, children should look out a window or into the distance for at least 20 seconds, researcher Dr. Caroline Klaver said.
Children should also be getting two hours of time outside every day.
Scientists Write ‘Hello World’ in Bacterial DNA With Electricity and CRISPR
The new system shows that it’s possible to encode information directly into bacteria
By storing data in bacteria like E. coli, the data is protected by the same machinery that the cell uses to protect its own DNA. (Janice Haney Carr/CDC)
One of the first things a budding software engineer learns is how to teach the computer to return the phrase, “Hello world!” So when experimenting with a new way to encode information into bacteria, that was the first message that scientists studying DNA data storage tried out.
A study published on January 11 in the journal Nature Chemical Biologydetails how the researchers led by Columbia University systems biologist Harris Wang used electricity and the DNA editing tool CRISPR to write “hello world!” into living bacteria’s DNA.
Genetic material like DNA is a potentially useful way to store data because it can store a lot of information in a small space. (For instance, ten full-length digital movies in the space of a grain of salt, Robert F. Service reports for Sciencemagazine.) Because DNA is integral to biology and bioengineering, the storage technology is unlikely to become outdated, John Timmer reports for Ars Technica.
“They are a very long way from having a working system that replaces our digital devices,” says EMBL-European Bioinformatics Institute senior scientist Nick Goldman to New Scientist’s Layla Liverpool. “But it’s a little step along the way to something that might do that.”
Computer code comes down to long strings of ones and zeros, and each digit is called a “bit” of information. A strand of DNA is a chain of four basic chemicals—abbreviated as A, C, G and T—that can be edited using bioengineering tools like CRISPR. In the new study, a change to a genetic sequence translated to a “one,” while no change translated to a “zero.” Any combination of six bacterial bits referred to a letter, number, space or punctuation mark, so the researchers called it a “byte.” (In a computer, a byte is made of eight bits.)
Using bacterial bytes, the scientists created the 12-character message “hello world!”
The electrical editing technique used in the new study builds on previous work led by Wang. In a 2017 study, researchers showed that they could make bacteria that use CRISPR to create a note in their DNA when they encounter the sugar fructose, per Science. The sugar sparked a series of events in each bacterium. First, the cell created a bunch of small rings of DNA, and that prompted CRISPR to snip the rings and save them in the bacterium’s own DNA.
In the new study, the researchers swapped sugar for electricity. Electricity changes one of the chemicals floating in the solution around the bacteria. The bacteria can sense the chemical change and set off a similar chain of events, resulting in a new chunk of code inserted into their DNA. By flipping the electricity on and off, the scientists could change the bacterial DNA code in precise locations.
The entire “hello world!” message didn’t fit in a single bacterium’s genome. Instead, the researchers created eight varieties of bacteria with three bits of information each. Pairs of bacteria strains provided the six bits necessary to create one letter or character. With 12 pairs of bacteria strains, each labeled with the order they should be read in, the scientists created the well-known message.
The system is still in its early days, Wang tells Science magazine. “We’re not going to compete with the current memory storage systems,” he adds.
But with more research, DNA data storage could have several benefits, Wang says. For one thing, genetic code is unlikely to become an outdated storage technology—and research is underway to make it ever-easier to edit and read DNA. Plus, writing data directly into living bacteria means the DNA is protected by organism and that the data will be copied into each new bacterium as the cells divide.
Wang says DNA inside of living bacteria could be a stable way to store data for medium to long-term storage.
“What you’re offering by putting it inside the cell is that the DNA is protected by the cell and the machinery that the cell has to protect its DNA,” says Wang to New Scientist.
The researchers even mixed their batches of data-carrying bacteria with potting soil and then recovered the message after a week. They estimate that the bacteria could hold onto their data for about 80 generations, per Ars Technica. However, the longer the bacteria spend copying their DNA, the more chance they have to introduce a mistake into the bits of information. That could confuse the message.
Harvard University biological engineer George Church tells New Scientist, “This field is progressing exponentially and this paper is a great example.”
Optical chaotic system is a central research topic due to its scientific importance and practical relevance in key photonic applications such as laser optics and optical communication. Because of the ultrafast propagation of light, all previous studies on optical chaos are based on either static imaging or spectral measurement, which shows only time-averaged phenomena. The ability to reveal real-time optical chaotic dynamics and, hence, control its behavior is critical to the further understanding and engineering of these systems. Here, we report a real-time spatial-temporal imaging of an optical chaotic system, using compressed ultrafast photography. The time evolution of the system’s phase map is imaged without repeating measurement. We also demonstrate the ability to simultaneously control and monitor optical chaotic systems in real time. Our work introduces a new angle to the study of nonrepeatable optical chaos, paving the way for fully understanding and using chaotic systems in various disciplines.
SIGN UP FOR THE SCIENCE ADVANCES eTOC
Get the latest issue of Science Advances delivered right to you!
Email
INTRODUCTION
Chaotic behavior is ubiquitous in nature. It has wide and profound influence on many disciplines ranging from fundamental sciences including biology, physics, and mathematics to applications including communication, cryptography, and robotics (1–7). Optical systems have been proven promising for studying chaotic behavior (8–11). Different mechanisms including laser instability (11–13), Kerr nonlinearity (9), and irregular cavities (14, 15) have shown strong chaos. The understanding of these chaotic phenomena in different optical systems is critical to both preventing chaos, when system stability is needed (16), and engineering chaos, when system performance is desired (3, 11, 12). Until now, the study of optical chaotic systems still relies on static imaging (17) and spectral measurement (11, 18). Consequently, only time-averaged effects are revealed, missing critical information about dynamic evolution and sensitivity of optical chaos. The real-time recording of optical chaotic systems has been hindered by the ultrafast movement of photons. Exposure times below picoseconds or imaging speeds above billion frames per second are required. Despite great improvements in the state-of-the-art electronic sensors, these speeds are beyond the capability of current complementary metal-oxide semiconductor and charge-coupled device (CCD) imaging technologies, due to the limited on-chip storage capacity and slow electronic readout speeds (19, 20). Other ultrafast imaging techniques such as Kerr gating have also been developed; however, they normally require repeated measurements under the condition that the chaotic events are highly repeatable (21, 22). The required precise repeatability is in great contradiction to the essence of chaos, which is ultrasensitive to initial conditions and infinitesimal fluctuations, thus nonrepeatable. The lack of ultrafast single-shot detection also limits the capability to control chaotic optical systems, which is highly desired for real applications.
In this study, by reporting a single-shot real-time recording of optical chaotic systems, we demonstrate a possible way to control optical chaos and monitor its dynamics at the same time. Using our compressed ultrafast photography (CUP) technique (23) (Fig. 1A), snapshots of light propagation in two-dimensional (2D) irregular optical cavities are taken at a speed up to 1 billion frames/s. Phase maps of irregular optical cavities are directly measured, which reveals full information of the system. Furthermore, we demonstrate the ability to control and monitor optical chaotic systems in real time by combining the Kerr gate and CUP techniques.
Fig. 1Schematic of the CUP system and the half-mushroom cavity.(A) Schematic setup of the CUP system. Light motions in the half-mushroom cavity are recorded by the CUP system in real time. (B) Half-mushroom cavity built from reflective mirrors. The definitions of s and p (= sinθ) are illustrated. (C) Simulated light trajectories in the regular mode. (D) The simulated Poincaré surface of section (SOS) phase space of the half-mushroom cavity showing both the regular modes (red lines) and the chaotic modes (blue dots). (E) Light trajectories in the chaotic mode.
RESULTS
We first study a 2D half-mushroom cavity, which is a typical cavity structure used in both theoretical and steady-state experimental studies of optical chaos phenomenon (24–27). Optical chaotic systems are built on the basis of the classical billiard chaos theory. Light propagation in closed linear 2D cavities with special boundary configurations can show chaotic properties. To characterize the light propagation dynamics, femtosecond laser pulses with 100-fs pulse duration and 800-nm center wavelength from a Ti-sapphire laser are fed into the 2D half-mushroom cavity at a grazing angle (Fig. 1A). The cavity is placed on the imaging plane of the CUP system (23). The CUP system is triggered by the Ti-sapphire laser to record the light propagation in the cavity. The total recording time is typically several thousand picoseconds, and the temporal resolution is 10 ps. Weak optical scattering is introduced so the CUP camera can capture motion of light (see Materials and Methods for details).
The half-mushroom cavity, shown in Fig. 1B and fig. S1, is characterized by the radius r of the quarter circle, the foot width w, and foot height h (Fig. 1C). The overall interior surfaces are light reflective and define the half-mushroom cavity boundaries for light propagation. Light propagation inside the cavities can be represented by the reflection position and angle on the quarter circle, which are the Birkhoff coordinates of optical chaotic systems (Fig. 1B). By plotting the Birkhoff coordinates, the Poincaré surface of section (SOS) can be constructed to characterize all system features in phase space (see section S2). In Fig. 1D, we show the simulated SOS of a half-mushroom cavity with r = 2 arbitrary units (a.u.), h = 0.5 a.u., and w = 1.2 a.u. (also see section S3.1). The SOS of the half-mushroom cavity is a mixed phase space, which distinctively shows two regimes: regular and chaotic regimes. In the regular regime, the trajectory of light propagation (Fig. 1C) has a constant reflection angle, showing a horizontal line in SOS. In the chaotic regime, the trajectory of light propagation (Fig. 1E) is ergodic and shows an exponential dependence on initial conditions: incident position described by s and incident angle described by θ (Fig. 1D).
To evaluate how the geometry of the cavity affects and controls its chaotic behavior, a standard half-mushroom cavity (Fig. 2A) and a deformed half-mushroom cavity (Fig. 2B) are built with the only difference being the tilted angle. One trajectory of light propagation inside the standard half-mushroom cavity is shown as time-lapse frames in Fig. 2C and in movie S1. The complete SOS phase map of the half-mushroom cavities is recorded by imaging light propagations under different light incident conditions with the CUP system. The SOS phase map helps us confirm CUP’s suitability for the study of optical chaos. After all light trajectories are recorded, the reflection positions and angles on the arc mirror are extracted to form the SOS phase map. For the standard half-mushroom cavity, the SOS phase map is shown in Fig. 2D. Compared to the simulated phase map of a perfect half-mushroom billiard shown in Fig. 1D (also see section S3.1), the sharp boundary between the regular and chaotic regimes disappears. Instead, we observe three types of light propagation in the SOS phase map: (i) Trajectories with ∣p∣ > 0.8 show small variances in ∣p∣ and are similar to those in the regular regime in the perfect half-mushroom cavity (Fig. 1D, red lines); (ii) stable periodic orbits with 0.5 < ∣p∣ < 0.8 are surrounded by invariant curves, which form small islands; and (iii) chaotic trajectories with ∣p∣ < 0.5 forming the chaotic sea are similar to those in the perfect half-mushroom cavity (Fig. 1D, blue dots). On the basis of the Kolmogorov-Arnol’d-Moser (KAM) theorem (28, 29), such an SOS phase map indicates that the geometry of the standard half-mushroom cavity used in the experiment deviates from the geometry of a perfect half-mushroom cavity (see section S3.2 for the simulated phase map of a half-mushroom cavity with minor deformations). This observation is further confirmed in the deformed half-mushroom cavity by intentionally tilting one mirror to a large angle (Fig. 2B). According to the KAM theorem, larger deformation leads to the disappearance of more invariant curves and to an increase in the chaotic regime. As shown in Fig. 2E, the regular trajectories with large ∣p∣ cannot be observed. The SOS phase map mainly consists of chaotic seas with small islands surrounded by invariant curves. The key features in the experimentally obtained phase map in Fig. 2E match well with those in the simulated phase map for this severely tilted half-mushroom cavity (see section S3.3). The above result reveals that cavity geometry is an essential parameter in controlling optical chaos.
Fig. 2Single-shot real-time imaging observation of controlling optical chaos by tuning geometry of optical cavities.As-constructed (A) regular half-mushroom and (B) tilted half-mushroom cavities built from reflective mirrors. (C) Time-lapse images showing the light trajectory inside the regular half-mushroom cavity (see corresponding video in movie S1). (D) Experimentally obtained Poincaré SOS phase map of the light modes inside the regular half-mushroom cavity in (A). (E) Experimentally obtained Poincaré SOS phase map of the light modes inside the tilted half-mushroom cavity in (B).
Small perturbation in a system can lead to markedly different results, which is the iconic feature of chaotic behavior. In the standard half-mushroom cavity (Fig. 2A), two light trajectories are recorded consecutively with a 1-ms time interval under the same experimental condition. Two such trajectories are shown as time-lapse frames in Fig. 3 (A and B, respectively) (see movie S2). Ideally, these two trajectories should be exactly the same due to the same experimental conditions. Experimental results from Fig. 3 (A and B) show that the two trajectories nearly coincide at the beginning but start to diverge substantially after 900 ps. Time evolution of two trajectories is plotted in SOS phase space in Fig. 3C, which clearly reveals the divergence of two trajectories as time elapses. Figure 3D shows how these two trajectories propagate in the phase space. Although the same experiment conditions are applied to the two light trajectories, the large divergence is due to the infinitesimal drift of the experiment setup over 1 ms. The sensitivity to the initial condition clearly indicates the chaotic behavior of the light propagation. In chaos theory, this behavior belongs to the category of deterministic chaos (30, 31), where, due to the extreme sensitivity of the system to the initial condition, small fluctuations in the initial condition make it impossible to predict long-term behavior in general. Any environmental disturbance may drift the experimental setup sufficiently to make the experiment nonrepeatable. This reveals that controlling small perturbation to chaotic system is also key factor to control optical chaos. Comparing to traditional simulation methods or steady-state experimental methods that cannot capture the sensitivity of optical chaos, the unique “one-shot” advantage of CUP becomes critical to studying the chaotic behavior.
Fig. 3Single-shot real-time imaging of two light trajectories under the same initial incident conditions.(A) Time-lapse images showing the light trajectory inside the half-mushroom cavity. (B) Second light trajectory under the same initial incident conditions as that in (A). The corresponding video data showing the temporal evolution of the light pathways in (A) and (B) are provided in the left and middle panels of movie S2, respectively. The right panel of movie S2 shows the combined movie overlaying both light pathways. Infinitesimal differences under the initial condition due to system shift propagates into vastly different light paths as compared to (A). (C) Evolution of the light paths in the Poincaré SOS phase spaces over time for the light trajectories in (A) and (B). (D) Poincaré SOS phase space shows full trajectories of the two lights.
We further developed the technique to control and monitor light propagation simultaneously in real time. It is realized with a special cavity design shown in Fig. 4A, which is a quarter Bunimovich stadium (32, 33). A successive single-shot light trajectory in Bunimovich stadium is shown in Fig. 4 (A to C) (see the corresponding video data in movie S3). A Kerr gate, consisting of a thin Bismuth Germanate (BGO) crystal and a plate polarizing beam splitter (PBS), is placed at the boundary between the rectangle and quarter-circle parts of the cavity. A Kerr gate can switch the light between regular and chaotic mode. When the Kerr gate is open, light propagation in the quarter Bunimovich stadium always shows chaotic behavior (32, 33). When the Kerr gate is closed, light pathways in both the rectangular and quarter-circle cavities separated by the Kerr gate are always nonchaotic (see sections S3 and S4) (29). The femtosecond laser at 800-nm center wavelength is first sent into a bulk lithium niobate crystal for frequency doubling to 400-nm center wavelength with 10% efficiency. A dichroic mirror is used to separate the 400- and 800-nm optical pulses. The 400-nm pulses are used to probe the light propagation dynamics in the cavity, and the 800-nm pulses are used to control the Kerr gate. A short-pass filter is placed before the CUP system to eliminate the scattered 800-nm light. The polarization of 400-nm probe light is adjusted to be s-polarized, so that light is reflected by the PBS. Therefore, light is confined in the rectangular cavity instead of the quarter Bunimovich stadium (Fig. 4A). The trajectory of light propagation is an invariant curve with constant reflection angles (Fig. 4A). With the 800-nm control light, the 400-nm probe light is changed to p-polarization, thus transmitting the PBS (from t = 380 ps in Fig. 4B). The cavity is changed from a rectangle to a quarter Bunimovich stadium, and the light propagation follows a chaotic trajectory instead of the original regular trajectory (Fig. 4B). At t = 470 ps, the Kerr gate is activated again to change the polarization of the 400-nm probe light back to s-polarization (Fig. 4C). Therefore, the probe light is confined in the rectangular cavity again (Fig. 4C). However, the light now propagates in a different regular mode instead of the original regular mode when t < 380 ps. This is also shown in the SOS phase space (Fig. 4D). If the Kerr gate never opened, then the light would follow the imaginary trajectory (Fig. 4D, light cyan dots). Instead, because of the switching of the Kerr gate, light trajectory transitioned to the chaotic mode at 390 ps (purple dots) and then transitioned back to a different regular mode after 470 ps (cyan dots).
Fig. 4Real-time control of regular and chaotic optical modes in the quarter Bunimovich stadium using a Kerr gate.Using a Kerr gate and a second control light pulse, the light modes can be controlled in real time to transition between regular modes and chaotic modes. Schematic and successive real-time recording of light trajectories inside the quarter Bunimovich stadium when (A) the Kerr gate remains closed, (B) the Kerr gate remains open from 380 to 470 ps, and (C) the Kerr gate remains closed. The corresponding video data showing the temporal evolution of the light pathway in this measurement is provided in movie S3. (D) The definitions of s and p (= sinθ) and the experimentally obtained Poincaré SOS phase map corresponding to the light dynamics in (A) to (C) are shown. The regular mode (cyan dot), chaotic mode (purple dot), and predictions of light trajectories if the Kerr gate never opens (light cyan dot) are shown.
DISCUSSION
By comparing with traditional time-integrating imaging methods, CUP has great advantages in studying chaos in optical cavities, providing more insights into optical chaos. As chaotic light traverses all spatial points inside the cavity (25, 29), a long-time exposure would overlap the images of the light paths, making it challenging to sort out individual light paths precisely, not to mention quantifying the time sequence of these light paths. The true value of CUP lies in its capability to obtain complete temporal information along with spatial information. One prominent example is to extract the distribution of the Poincaré recurrence time (34), which is critical for studying many chaotic cavities (35, 36). In addition, the temporal SOS phase map can reveal the real property of optical cavities if compared with the traditional SOS phase map. It has been found that the traditional SOS phase map may generate wrong light survival probability in a leaky chaotic limaçon-like cavity as a function of the physical time if compared with the result from a true-time SOS phase map, because a traditional SOS phase map may associate each collision with the same time and may overestimate the collisions that happened within a short period (37). Another example is the dynamic tunneling between regular modes and chaotic modes (27, 38), which can possibly be directly visualized by CUP. It facilitates the study of the dynamic tunneling effect in chaotic cavities and helps in engineering the applications using the tunneling effect (39).
The capability to directly observe and control the optical chaotic behavior in real time in both the spatial and temporal domains opens the door to new research paradigms for optical chaotic systems beyond the traditional theoretical and static experimental approaches. The first-generation CUP system operates at an imaging speed of 100 billion frames/s, with the ability to resolve light dynamics in centimeter-sized optical cavities. CUP systems with an imaging speed of 10 trillion frames/s have already been demonstrated (40), which may allow the ability to resolve light dynamics in integrated nanophotonic cavities. Moreover, by integrating diffraction gratings into CUP systems, the spectral information can also be obtained, which is critical for chaotic systems induced by nonlinear optical processes.
MATERIALS AND METHODS
CUP systems
The operation of the CUP imaging system can be divided into two operational steps: (i) the real-time image acquisition and (ii) the subsequent image reconstruction. In the first step of the measurement, each 2D image frame of the input object video is first projected onto a digital micromirror device (DMD), which encrypts each image with a 2D pseudorandom binary pattern. The encrypted image is then projected onto a stream camera with a widened entrance slit. Within the stream camera, the image is temporally sheared by a sweeping electric field. The resulting sheared image is temporally integrated by a CCD detector array. In the image reconstruction step, the captured data are used to reconstruct the object video based on the encrypted information using compressive sensing algorithms.
The experimental setup is shown schematically in Fig. 1A. The optical cavity is placed at the object plane of the CUP system. To image light propagation, water vapor is used to scatter light into the CUP system. As the streak camera is highly sensitive (single-photon sensitivity in principle), only weak scattering is needed. Therefore, the effects to chaos from these scattering events can be neglected compared with the effects from the deformation of the cavities and opening/closing Kerr gates. The light propagation is first imaged by a 4f system consisting of two lenses with focal lengths of 150 and 25.4 mm, respectively. The intermediate image is then passed to a DMD by another 4f imaging system consisting of a tube lens (focal length, 150 mm) and a microscope objective (focal length, 50 mm; numerical aperture, 0.16). To encode the input image, a pseudorandom binary pattern is generated and displayed on the DMD, with a binned pixel size of 21.6 μm by 21.6 μm (3 3 3 binning). The light reflected from the DMD is collected by the same microscope objective and another tube lens with a focal length of 200 mm and imaged onto the entrance slit of the streak camera. To allow 2D imaging, this entrance slit is opened to its maximal width (~5 mm). Inside the streak camera, a sweeping voltage is applied, deflecting the encoded image frames according to their times of arrival. The final temporally dispersed image is captured by a CCD with a single exposure.
This is an open-access article distributed under the terms of the Creative Commons Attribution-NonCommercial license, which permits use, distribution, and reproduction in any medium, so long as the resultant use is not for commercial advantage and provided the original work is properly cited.
, Ballistic 2-D imaging through scattering walls using an ultrafast optical Kerr gate. Science 253, 769–771 (1991).Abstract/FREE Full TextGoogle Scholar
Acknowledgments: Funding: X.Y. and H.W. would like to acknowledge the support from the Army Research Office Young Investigator Program (grant no. W911NF-18-1-0268), the Air Force Office of Scientific Research (FA9550-15-1-0514), and the NSF (grant no. ECCS-1653870). L.V.W. acknowledges the support by the NIH grant DP1 EB016986 (NIH Director’s Pioneer Award) and the NIH grant R01 EB028277. Author contributions:H.W. and L.F. conceived the research project. L.F. and X.Y. constructed the optical cavities and performed the experiments. X.Y. carried out the simulations of the classical billiard optical chaos. L.V.W. supervised the development of the CUP method and the CUP measurements and envisioned applications in chaotic phenomena such as optical rogue waves. H.W. and L.V.W supervised the overall research effort. All authors discussed the results and co-wrote the manuscript. Competing interests: The authors declare that they have no competing interests. Data and materials availability: All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials. Additional data related to this paper may be requested from the authors.
For the first time, scientists have measured the different types of genomic DNA changes that occur in skin cells, finding that mutations from ultraviolet (UV) light is especially common, but Black individuals have lower levels of UV damage compared to white people. Dmitry Gordenin and colleagues at the National Institute of Environmental Health Sciences, report these findings January 14 in PLOS Genetics.
The DNA in our skin cells suffer damage from sources inside and outside the body, leading to genomic changes such as mutations that may lead to cancer. UV light is the major source of these mutations, but byproducts of cellular metabolism, like free radicals, and DNA copying errors that occur during cell division also cause genomic changes. These mutation-causing mechanisms are well known, but previously, no one had been able to accurately measure the relative contributions from each source.
In their new paper, Gordenin and his colleagues quantified the amounts of each type of genomic changes by sequencing the genomes of skin cells donated from 21 Black and white individuals, ranging in age from 25 to 79. The researchers discovered that the total amount of genomic changes from metabolic byproducts accumulates as a person gets older, while the amount of genomic changes caused by UV damage is unrelated to a person’s age. Additionally, they showed that genomic changes from UV light is common, even in skin cells typically shielded from the sun, but it was less prevalent in Black donors compared to white donors.
The researchers suspect that Black individuals may be better protected from UV light due to having higher levels of the skin pigment melanin. Supporting this idea, is the fact that Black people have much lower rates of skin cancer compared to white people. Overall, the new study provides an accurate estimate of the genomic changes that occur in skin cells due to different types of DNA damage, and establishes the normal range of somatic genomic changes across a wide range of ages and of different races, providing a baseline for future research.
The authors add, “The new study provides an accurate estimate of the genomic changes that occur in skin cells due to different types of DNA damage, and establishes the normal range of somatic genomic changes across a wide range of ages and of different races, providing a baseline for future research.”
More information: Saini N, Giacobone CK, Klimczak LJ, Papas BN, Burkholder AB, Li J-L, et al. (2021) UV-exposure, endogenous DNA damage, and DNA replication errors shape the spectra of genome changes in human skin. PLoS Genet 17(1): e1009302. journals.plos.org/plosgenetics … journal.pgen.1009302Journal information:PLoS GeneticsProvided by Public Library of Science
Weekly DailySpecial delivery: Stem cells can be modified to produce a therapeutic protein in the brain.
Laguna Design / Science Photo Library
Two unpublished studies detail improved methods for delivering gene therapies to the brain: One involves a type of stem cell that can produce gene-altering proteins on-site; the other taps an engineered virus to target neurons efficiently and noninvasively.
One of the biggest hurdles for targeted gene therapy is getting enough treatment to the right spot. In the first study, researchers overcame this obstacle by developing stem cells that produce a therapeutic protein inside the brain.
The team is using the approach to develop a treatment for Angelman syndrome, which is caused by mutation in or deletion of the maternal copy of the gene UBE3A. Because the paternal copy of the gene is typically silent, loss of the maternal copy results in an absence of UBE3A protein. People with Angelman syndrome usually have intellectual disability and motor impairments, and many are autistic.
The researchers had previously used modified stem cells to produce a protein that can activate the paternal copy of UBE3A. Transplanting the cells into the brains of Angelman syndrome model mice boosts levels of UBE3A protein, they found. However, the treatment required multiple direct injections into the animals’ brains.
In the new work, they instead tried injecting the cells into a pocket of cerebrospinal fluid at the base of the skull — an approach that is less invasive and can be performed multiple times. They compared the results with direct injection into the animals’ hippocampus. In both cases, the mice had UBE3A expression in the brain for up to three weeks.
Mice that received direct injection of the stem cells had fewer Angelman syndrome traits than controls, as measured by their motor skills.
This suggests that though the new route is effective, it may not provide a high enough dosage, says Peter Deng, a postdoctoral researcher in Kyle Fink‘s lab at the University of California, Davis, who presented the work. And because the transplanted cells produce protein for only a limited period of time, the effects are temporary — a limitation the team is addressing.
Deng and his colleagues also found that monkeys treated with the stem cells had the therapeutic protein throughout their brain and spinal cord three weeks after injection, which suggests the approach has potential for treating people.
Overcoming barriers:
The second approach presented at the conference improves the delivery of a more permanent form of gene therapy that uses adeno-associated viruses (AAVs).
Researchers typically inject these viruses directly into the brain, and the viruses usually only affect cells immediately surrounding the injection site.
“You’re required to use a ton of the virus” to penetrate the whole brain, says Jerzy Szablowski, assistant professor of neuroengineering at Rice University in Houston, Texas, who presented the work.
One potential workaround is to inject the AAV into the blood and use focused ultrasound to temporarily open up the blood-brain barrier, allowing the AAV to cross into the brain. Sometimes with this approach, however, the virus also inserts itself into other organs.
In their new work, the team developed AAVs that more easily cross the blood-brain barrier and more selectively target neurons than previous versions do. As a result, the new AAVs can be given in lower doses, reducing the amount of tissue affected outside the brain, Szablowski says.
To identify the most efficient AAV, Szablowski and his colleagues designed 2,100 new viruses, injected them all into the bloodstream of mice and applied focused ultrasound to the animals’ skulls. The mice had been engineered so that AAVs that successfully inserted themselves into a neuron got tagged with a marker. The team performed genomic sequencing on the mouse brains a few weeks later and read out the levels of viruses.
Compared with the previously most effective AAV, the top five newly identified AAVs targeted twice as many cells in the brain (including more neurons), and nearly half as many cells outside the brain, the researchers found.
The approach could be used to more efficiently deliver treatments for conditions such as Angelman syndrome or Parkinson’s disease, the team says.
Drones have been used to send quantum internet signals
Entangled photons have been sent between two drones hovering a kilometre apart, demonstrating technology that could form the building blocks of a quantum internet.
When a pair of photons are quantum entangled, you can instantly deduce the state of one by measuring the other, regardless of the distance separating them. This phenomenon, which Albert Einstein dismissively called “spooky action at a distance”, is the basis of quantum encryption – using entangled particles to ensure communications are secret.
Quantum networks are far more secure than the existing internet because any attempt to eavesdrop changes the state of the photons, alerting the recipient to foul play.
Entangled photons have been transported more than 1000 kilometres in tests between a satellite and ground stations before, but now Zhenda Xie at Nanjing University in China and his colleagues have shown that links can be made over shorter distances with relatively inexpensive hardware. It is also the first time that photon entanglement has been transmitted from one moving device to another.
A laser on board one of the 35-kilogram drones created a pair of entangled photons by splitting a single photon with a crystal. One photon was sent directly to a ground station and the other to a second drone a kilometre away via a relay drone.
Motorised devices on each drone moved to ensure that the receivers and transmitters always lined up, and photons were focused and steered through the relay drone by a short piece of fibre-optic cable. The state of each photon was measured at the ground station and the results proved that the photons remained entangled.
Xie hopes that connections of over 300 kilometres can be achieved by more advanced drones at high altitude, free of the distorting influence of pollution and weather, and that smaller, more cost-effective drones could be produced for local connections, perhaps even to moving vehicles. All of these devices could link to satellites for global transmission.
The achievement marks an important step towards a quantum internet, says Siddarth Joshi at the University of Bristol, UK. He agrees that drones could become the final chain in links from one part of the world to another, such as from your local relay station to your home or vehicle. “You’re driving around in your car and you want to maintain secure quantum communications, so you have these drones flying around behind you,” he says.
Myungshik Kim at Imperial College London believes that engineering such complex optics into moving drones, especially given that small rotational differences can make it extremely difficult to maintain quantum connections, represents a strong technical advance.
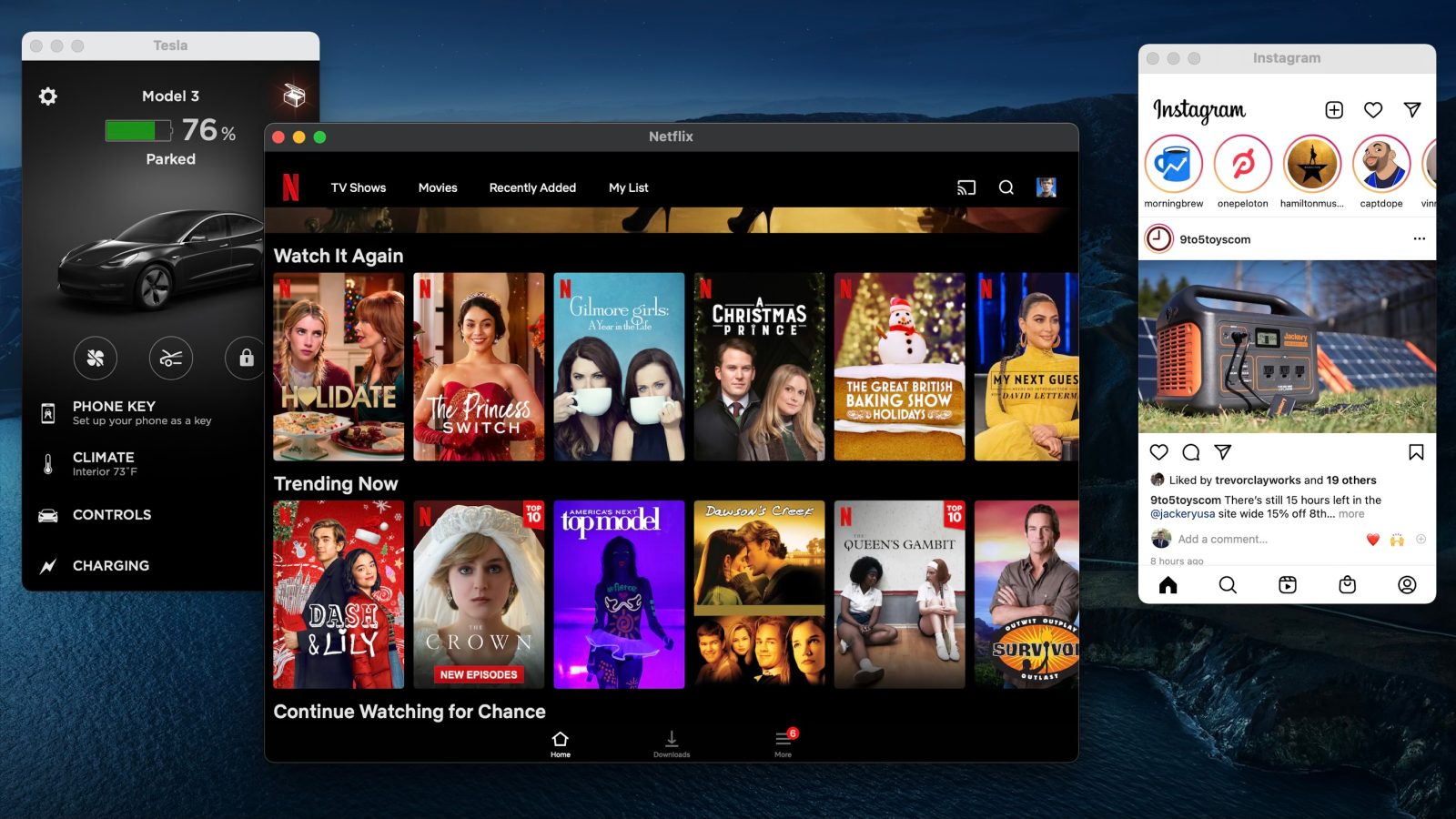
Earlier this week, 9to5Mac reported that Apple would soon start blocking users from side loading iPhone and iPad applications to their M1 Macs. Now, Apple has officially flipped the server-side switch to implement this change.
This means that it is no longer possible to use an app such as iMazing to side load unsupported applications from your iPhone or iPad to your M1 Mac.
As a refresher, Apple Silicon Macs allow users to run iOS and iPad applications on their Mac, but developers can opt out of allowing their apps to be installed on the Mac. This is the path that many developers have taken, making the necessary change in App Store Connect to remove their app from the Mac App Store.
But with that being said, until today, you could manually install iOS apps like Netflix, Instagram, and Facebook on an M1 Mac by using their respective IPA files downloaded under a valid Apple ID. Many people were using tools such as iMazing to complete this process.
9to5Mac has now confirmed that, starting today, this is no longer possible unless the application is available on the Mac App Store. Apple has flipped the necessary sever-side switch to block iPhone and iPad applications from being installed on Apple Silicon Macs.
The change applies to M1 Macs running macOS Big Sur 11.1 as well as the developer or public beta of macOS Big Sur 11.2. The only difference is that users running the macOS Big Sur 11.2 beta will see a more specific error message.
Here is the error you’ll see when you try to install an unsupported iPhone or iPad app on your M1 Mac running macOS 11.2 beta:
If you attempt to side load an iPhone or iPad app on an M1 Mac running macOS 11.1 Big Sur, you’ll see an error saying that the app installation was unsuccessful and to “try again later.”
If you already side loaded an application from your iPhone or iPad to your M1 Mac, it will still run properly. This change only affects new application installations. Additionally, if you had previously downloaded an .IPA file, you can still install it.
The change itself was made to the App Store system that delivers the actual .IPA file and it is all part of Apple’s APIs that manage the DRM (Digital Rights Management) protections of the operating system. Because of this, it’s unlikely that a workaround will present itself in the future.
FTC: We use income earning auto affiliate links.More.
Harvard scientists have created a swarm of swimming robots that could pave the way for new methods of environmental monitoring and searching coral reefs.
The autonomous “Bluebots” synchronize their movements like a real school of fish, basing their individual decisions on the behavior of their neighbors.
The collective Blueswarm deploys a 3D vision-based coordination system and 3D locomotion to navigate, while each individual bot uses two cameras and LED lights to follow its schoolmates.
Their fisheye lens cameras detect the LEDs of nearby Bluebots, and use a novel algorithm to calculate where they are and where they’re going. This allows the school to independently organize its movements.
Study author Florian Berlinger said the system enables each robot to implicitly react to the behaviors of its neighbors:
If we want the robots to aggregate, then each Bluebot will calculate the position of each of its neighbors and move towards the center. If we want the robots to disperse, the Bluebots do the opposite. If we want them to swim as a school in a circle, they are programmed to follow lights directly in front of them in a clockwise direction.
The researchers tested out their Blueswarm on a simulated search mission using a red light in a tank.
The bots used their dispersion algorithm to spread across the tank until one detected the light. Its LEDs then started to flash, triggering the aggregation algorithm in the rest of the swarm to pull the entire school around the signaler.
Study author Florian Berlinger said the Blueswarm could be highly useful in areas that are inaccessible or dangerous to humans:
In these situations, it really benefits you to have a highly autonomous robot swarm that is self-sufficient. By using implicit rules and 3D visual perception, we were able to create a system that has a high degree of autonomy and flexibility underwater where things like GPS and WiFi are not accessible.
The team now plans to use their research to develop further underwater swarms that can perform environmental monitoring and inspect coral reefs or man-made underwater structures. They could even unearth new insights about the real fish that inspired them.
You can read their research paper in the journal Science Robotics.
Cancer cell during cell division. Credit: National Institutes of Health
Our biological or circadian clock synchronizes all our bodily processes to the natural rhythms of light and dark. It’s no wonder then that disrupting the clock can wreak havoc on our body. In fact, studies have shown that when circadian rhythms are disturbed through sleep deprivation, jet lag, or shift work, there is an increased incidence of some cancers including prostate cancer, which is the second leading cause of cancer death for men in the U.S. With an urgent need to develop novel therapeutic targets for prostate cancer, researchers at the Sidney Kimmel Cancer—Jefferson Health (SKCC) explored the circadian clock and found an unexpected role for the clock gene CRY-1 in cancer progression. The study was published on January 15th in Nature Communications.
“When we analyzed human cancer data, the circadian factor CRY-1 was found to increase in late stage prostate cancers, and is strongly associated with poor outcomes,” explains Karen Knudsen, MBA Ph.D., executive vice president of oncology services for Jefferson Health and enterprise director of SKCC, and senior author of the study. “However, the role CRY-1 in human cancers has not been explored.”
A common therapy for prostate cancer involves suppressing the male hormone androgen and/or the androgen receptor, as prostate tumors require androgens to develop and progress to advanced disease. With their collaborators in the U.S. and Europe, the researchers found that CRY-1 is induced by the androgen receptor in prostate tumor tissue obtained from patients, thus explaining in part the high levels of CRY-1 observed in human disease.
“This was a clear indication of CRY-1’s link to prostate cancer,” says Ayesha Shafi, Ph.D., a postdoctoral researcher in Dr. Knudsen’s lab and first author of the study. “As we looked further into the role of CRY1, we unexpectedly found that the circadian factor was altering the way that cancer cells repair DNA.”
Cancer treatments aim to damage the DNA in cancer cells and cause defects in repair mechanisms; eventually the cells self-destruct when the damage is severe. The researchers probed CRY-1’s possible role in DNA repair in cultured cells, animal models and tissue harvested from prostate cancer patients. They first induced DNA damage by exposing cancer cells to radiation and found that CRY-1 levels became elevated, indicating that it was responding to this type of damage. They also found that CRY-1 directly regulates the availability of factors essential for the DNA repair process, and alters the means by which cancer cells respond to DNA damage. The findings suggest that CRY-1 may offer a protective effect against damaging therapies.
“The fact that CRY-1 is elevated in late-stage prostate cancer may explain why androgen-targeting treatments become ineffective at those later stages,” says Dr. Shafi. “It also tells us that if a tumor has high levels of CRY-1, DNA repair targeting treatments may be less effective for them.”
“Not only have we outlined a role for CRY-1 outside of its canonical function in circadian rhythms, Dr. Shafi’s findings are the first to reveal the means by which CRY1 contributes to aggressive disease,” adds Dr. Knudsen. “It’s notable that the pro-tumor functions of CRY1 may be viable targets to treat prostate cancer, and this is a direction that Dr. Shafi’s future work will explore.”
Looking ahead, the team plans to explore how best to target and block CRY-1 and what other existing therapies may work synergistically to hinder DNA repair in prostate cancer cells. They also plan to study more circadian rhythm genes and determine how circadian disruption may affect cancer treatment.
“It’s been shown that circadian disruptions can affect efficacy of treatment, but also that aligning treatment with the body’s natural rhythms or giving therapy at certain times of the day can be beneficial,” explains Dr. Knudsen. “Our findings open up a multitude of important research questions exploring the link between the circadian clock and cancer.”
More information: “The circadian cryptochrome, CRY1, is a pro-tumorigenic factor that rhythmically modulates DNA repair,” Nature Communications (2021). DOI: 10.1038/s41467-020-20513-5Journal information:Nature Communications
Besides the intended differences, web browsers based on Chromium offer an underlying experience that’s mostly identical to Chrome. Google recently discovered that users of third-party Chromium browsers have inadvertently been able to access data and other sync features reserved for Chrome.
“Some” Chromium browsers today can leverage features and APIs that are “only intended for Google’s use.” This includes Click to Call and, notably, Chrome Sync. The latter is responsible for syncing bookmarks, extensions, history, settings, and more across signed-in devices running the first-party browser.
This meant that a small fraction of users could sign into their Google Account and store their personal Chrome sync data, such as bookmarks, not just with Google Chrome, but also with some third-party Chromium based browsers.
As a result, users logged into Google sites on Chromium browsers are able to see their old bookmarks and other data from previous Chrome usage.
This inadvertent access was discovered during a recent audit and Google will be “limiting access to [its] private Chrome APIs” from March 15th.
Guidance for vendors of third-party Chromium based products is available on the Chromium wiki.
Users that have been benefiting from this accidental integration — to keep bookmarks in sync — will not lose any information. Any Chrome data stored locally will remain available, while it’s also still in your Google Account. Meanwhile, Chrome bookmarks remain transferable and can be easily exported to a new browser through existing methods.

/https://public-media.si-cdn.com/filer/d6/b4/d6b4dab9-b80c-4c31-b46d-465460bcb58f/10068_lores.jpg)




 Special delivery: Stem cells can be modified to produce a therapeutic protein in the brain.
Special delivery: Stem cells can be modified to produce a therapeutic protein in the brain.